La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 1
LE POTENTIEL DE DÉCOUVERTE ARCHÉOLOGIQUE DANS LES TERRAINS ORGANIQUES DU CANADA
Jacques Cinq-Mars
Conservateur, Archéologie du Québec
Commission archéologique du Canada
Musée canadien des civilisations
AVANT-PROPOS
Cet article a été publié originalement dans la Revista de Arqueología Americana, no 1, pp. 9-32, 1990. Les changements qui ont été apportés à cette version électronique consistent en quelques corrections mineures qui n’affectent en rien le contenu initial et en une présentation rafraîchie des cinq illustrations (Figures 1 – 5) et de la liste faunique (Tableau 1).
Pour ce qui est des références , on peut noter que l’article de Cinq-Mars et Morlan 1989, mentionné dans le texte, a finalement été publié:
Cinq-Mars, Jacques and Richard E. Morlan. 1999. Bluefish Caves and Old Crow Basin: A New Rapport, in Ice Age Peoples of North America, ed. by Robson Bonnichsen and Karen L. Turnmire, pp. 200-212. Corvallis: Center for the Study of the First Americans, Oregon State University Press.
Par ailleurs, celui de Cinq-Mars et Nelson 1989 n’a malheureusement pas encore vu le jour.
Finalement, des détails additionnels concernant les datations disponibles (brièvement mentionnées dans le texte) peuvent être obtenus en consultant la Banque des Datations par le Radiocarbone en Archéologie canadienne (BDRAC) à:
www.canadianarchaeology.com/radiocarbon/card/card.htm
Jacques Cinq-Mars, Hull, février 2001.
INTRODUCTION
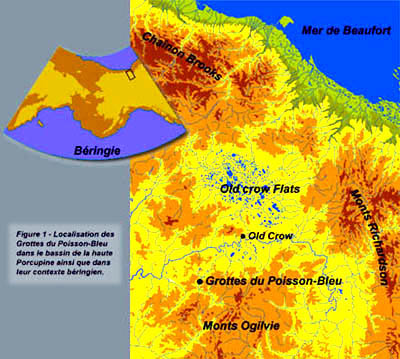
En réponse au besoin qu’avait son époque d’expliquer les réalités autochtones du Nouveau-Monde, José de Acosta imaginait, il y a 400 ans, ce qui allait devenir le détroit de Béring, c’est-à-dire, un lieu où une proximité intercontinentale relative aurait été propice aux mouvements de populations entre l’Asie et l’Amérique. Cet endroit forme encore aujourd’hui, avec les territoires adjacents (Fig. 1), le canevas géographique sur lequel ont été tissées de nombreuses hypothèses plus ou moins viables, variations portant sur le thème du peuplement des Amériques. Suite à de nombreuses études récentes et dans un effort de synthèse biogéographique, ces régions en sont venues à être connues sous le nom de Béringie (Hopkins 1959, 1967; Hopkins et al. 1982; Hultén 1968). Plus précisément, pour notre propos, nous ferons référence à la Béringie orientale, territoire correspondant à ces parties de l’Alaska et du Yukon qui, sauf dans quelques zones alpines, ne connurent pas les avances glaciaires du Pléistocène.
Maintes fois stimulées par les besoins propres à l’archéologie, les recherches interdisciplinaires effectuées dans ces régions, au cours des dernières décennies, ont rendu possible une appréhension plus juste des dimensions paléo-géographiques du concept de Béringie. Elles ont aussi permis de mieux cerner divers aspects de l’évolution d’un milieu pléistocène auquel durent s’adapter ces populations humaines dont certaines allaient, un jour, déborder plus au sud. Le concept de Béringie, ainsi que ses variantes et dérivés (Note 1) , peut donc être considéré comme étant essentiel à la logique interne des discours archéologiques qui, dans un cadre chronologique pléistocène, traitent des origines des premières populations autochtones du Nouveau-Monde. Cependant, il faut bien noter que les données pertinentes issues de ces travaux demeurent peu nombreuses et ce, surtout en raison des difficultés inhérentes à la pratique archéologique dans ces régions. En conséquence, il est opportun de clarifier quelques aspects de cette archéologie béringienne dont certaines des dimensions pour le moins hémisphériques, ont si souvent défrayé la chronique.
On peut diviser en deux groupes les faits archéologiques, ainsi que les interprétations les plus récentes et les plus raisonnables qu’on en a fait. Le premier intéresse avant tout le Tardiglaciaire et est constitué par un ensemble complexe de données qui proviennent d’un nombre grandissant de gisements découverts en Alaska et au Yukon. Sans être concluante, cette information semble de plus en plus en mesure d’apporter des solutions à la question du peuplement actuel des régions boréales et arctiques du Nord-Ouest ainsi qu’à celle des origines clovisiennes. Dans ce dernier cas, elle tend à satisfaire les tenants d’une série chronologique « courte ». L’autre ensemble de données est plus discret et, pour le moment, sans apparence de continuité avec le premier. Il résulte avant tout des travaux de recherche effectués au Yukon septentrional et dont les résultats suggèrent des manifestations culturelles plus anciennes, datant du Pléniglaciaire et même, comme nous allons le voir, de périodes antérieures. Ils amènent, quant à eux, de l’eau au moulin des tenants de séries chronologiques « longues ».
C’est à partir de certains de ces matériaux, provenant plus particulièrement des grottes du Poisson-Bleu (Bluefish Caves), au Yukon septentrional, que nous allons essayer d’éclairer d’une lumière béringienne quelques facettes de cette archéologie traitant des origines.
La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 2
DESCRIPTION DES GROTTES DU POISSON-BLEU
Localisation et environnement
Les grottes du Poisson-Bleu (Note 2) sont situées à 54 km au sud-ouest du village Vuntut Gwichin d’Old Crow, le long du cours moyen de la rivière Bluefish, un tributaire de la rivière Porcupine (Fig. 1). La région en est une de collines calcaires d’âge devonien (Norris 1985) formant l’avancée septentrionale du chaînon Keele, lui-même contrefort du grand massif des monts Ogilvie qui marque le centre du Yukon. Ces collines forment aussi la bordure sud-ouest d’un énorme réseau de bassins lacustres qui, au Pléistocène supérieur, furent ennoyés par les eaux des lacs glaciaires Bluefish, Old Crow et Bell (Hughes 1972). Le milieu est caractéristique de la forêt boréale en région montagneuse (Ritchie et al., 1982) avec peuplements d’épinettes (Picea glauca et Picea mariana) sur les pédiments,et avec zones de toundra plus ou moins continues, le long des crêtes de plus de 750 m d’altitude. Le climat est à l’avenant et l’ensemble du paysage est façonné par les multiples phénomènes périglaciaires propres aux régions à pergélisol continu.
Contexte géomorphologique

C’est à l’extrémité ouest d’une crête dominant un rétrécissement de la vallée de la Bluefish que se trouvent les grottes, nichées au pied d’une série d’affleurements calcaires qui surplombent la rivière d’environ 250 m. Elles sont au nombre de trois (grottes I, II et III) et consistent, en fait, en de petites cavités dont les dimensions volumétriques varient approximativement entre 10 et 30 m3 (Fig 2). Quant à leur genèse, il semble que, contrairement à ce que nous avions suggéré précédemment (Cinq-Mars 1979), elle s’inscrive dans un contexte géographique et chronologique beaucoup plus large, c’est-à-dire, celui du développement d’un karst régional, présentement à l’étude (Cinq-Mars et Lauriol 1985; Lauriol et al. 1989; Roberge et al. 1986). Autrement dit, il ne s’agirait pas de cavités ou porches de gélifraction mais plutôt de vestiges d’un ancien réseau karstique fortement réduit et mis au jour par l’érosion des versants.
Le remplissage
Les sondages et les fouilles effectués à l’intérieur aussi bien qu’à l’extérieur des trois grottes, ainsi que les analyses subséquentes des sédiments, ont révélé un mode de remplissage caractérisé par une uniformité relative des dépôts et de leur mise en place (Fig. 3). La lecture simplifiée peut s’en faire comme suit:
- à la base, une surface rocheuse (roche-mère) parsemée de fragments cryoclastiques avec, par endroits, dans quelques fissures et cuvettes, des plages résiduelles de sédiments très altérés;
- le sédiment sus-jacent, d’origine éolienne, est un lœss relativement homogène qui peut atteindre plus d’un mètre d’épaisseur et au sein duquel on retrouve, selon les endroits, des quantités variables d’éléments cryoclastiques provenant surtout, soit des parois et du plafond des cavités, soit des parois abruptes qui surplombent d’environ 10 m l’entrée des porches; les résultats d’analyses granulométriques et sédimentologiques effectuées sur des échantillons prélevés sous le larmier de la grotte II , nous permettent de suggèrer que, malgré son apparente homogénéité, ce lœss serait divisible en trois faciès qui réflètent les conditions de transport des matériaux constituants ainsi que leur lieu d’origine(Note 3);
- recouvrant le lœss de façon soit graduelle, soit discordante, nous trouvons, à l’extérieur des grottes, un humus à cailloutis (cryoclastique) dont l’épaisseur peut atteindre, par endroits, un peu plus d’un mètre; cette couche s’amenuise rapidement à l’entrée des porches pour se transformer, à l’intérieur, en un simple enrichissement organique de la surface du lœss;
- enfin, le remplissage est scellé par une végétation herbacée et arbustive à l’extérieur des grottes et, à l’intérieur, par un couvert discontinu de fougères, de mousses et de lichens.
- en premier lieu, notons un assemblage inférieur caractéristique d’une toundra riche en espèces herbacées; stratigraphiquement, cette zone correspond à la partie inférieure du lœss;
- lui succède, dans la partie supérieure du lœss, une zone dont le caractère principal consiste en une augmentation et une représentation maximale du bouleau (Betula);
- enfin, cette portion du remplissage que nous avons qualifié d’humus à cailloutis a fourni un assemblage caractérisé par une baisse sensible des espèces herbacées ainsi que par une augmentation importante des courbes de l’épinette (Picea) et de l’aulne (Alnus)

Cette présentation schématique du remplissage ne tient pas compte des traces nombreuses résultant de l’action des phénomènes périglaciaires tels que la cryoturbation et la gélifluxion, et qui en rendent parfois l’interprétation difficile. Nous nous trouvons donc en face de gisements qui se prêtent mal à une lecture stratigraphique fine et dont le décodage nécessite, peut-être plus que partout ailleurs, l’apport d’autres types de données.
Données palynologiques
L’analyse de sédiments provenant de l’intérieur de la grotte I et de l’extérieur de la grotte II a fourni des diagrammes polliniques d’où se dégage, malgré la nature des dépôts qu’ils servent à illustrer, une certaine intégrité. Ces diagrammes ont déjà fait l’objet de présentations (Cinq-Mars 1979; Ritchie et al. 1982) et nous nous contenterons, ici, d’en dégager les traits les plus signifiants:
Malgré la faiblesse de sa définition, cette biostratigraphie pollinique se compare bien à celles issues d’autres investigations palynologiques régionales (Ritchie et Cwynar 1982; Ritchie 1984). Ainsi, il est clair que la partie inférieure du lœss recèle les traces d’un milieu à toundra herbacée à caractères xérothermiques. À ces conditions environnementales très différentes de celles qui prédominent aujourd’hui, succèdent les échos d’une série de changements conduisant, d’abord, au développement d’une toundra arbustive à bouleau et, enfin, à l’établissement rapide d’une forêt boréale très proche de celle que nous connaissons.
Cette comparaison nous permet d’identifier quelques repères chronologiques utiles. Ainsi, il appert que le développement de la toundra arbustive, marqué par la croissance de la représentation du bouleau, date d’environ 14,000 – 13,500 ans BP et que l’apparition des conditions propres au développement de la forêt boréale à épinette se situe, dans notre région, autour de 10,000 ans BP. Enfin, soulignons que cette correspondance entre les diagrammes polliniques des gisements du Poisson-Bleu et les diagrammes régionaux sert à étayer la notion, mentionnée précédemment, d’intégrité relative du remplissage et ce, malgré l’inévitable brouillage périglaciaire.
Données paléontologiques
Les grottes ont, toutes les trois, fourni des milliers de vestiges osseux dont l’excellente conservation résulte de conditions sédimentologiques et taphonomiques très favorables. Cette faune est composée de grands et petits mammifères, incluant une série importante de microtinés, ainsi que des oiseaux et des poissons (Tabl. 1). Elle peut être divisée en deux séries provenant, respectivement, du lœss et de l’humus à cailloutis.
La faune du lœss est très riche, tant par sa quantité que par sa complexité et sa diversité. En ce sens, elle se conforme à ce que Guthrie (1982, 1985) a qualifié de « faune à mammouth » (Mammoth fauna) du Pléistocène supérieur béringien. Dans les gisements du Poisson-Bleu, cette dernière est surtout composée, pour ce qui est de la mégafaune, de formes telles que le cheval (Equus lambei), le caribou ou renne (Rangife r tarandus), le mouflon (Ovis dalli), le bison (Bison priscus), l’élan ou orignal (cf. Alces alces), le wapiti ou cerf élaphe (Cervus elaphus) et le mammouth (Mammuthus primigenius); elle contient également l’antilope saïga (Saiga tatarica), le bœuf musqué (Ovibos moschatus), l’ours (Ursus), le loup (Canis lupus) et le lion (Panthera).
Par contre, le matériel osseux provenant de l’humus à cailloutis possède, quantitativement aussi bien que qualitativement, des traits caractéristiques d’une mégafaune régionale fortement appauvrie, signes indubitables d’extinctions massives et d’extirpations régionales aussi bien que continentales. En fait, nous n’y avons recueilli que quelques spécimens épars de caribou et de mouflon, représentants discrets d’une mégafaune holocène régionale qui comprend aussi l’orignal, l’ours et le loup.
Comme dans le cas de la palynologie, nous nous retrouvons, au regard de la faune, face à un contraste qui s’accorde bien avec une césure chrono-stratigraphique (lœss/humus à cailloutis) évidente (Note 4). Sa signification, même si elle n’est pas encore vérifiée, semble reliée à des changements globaux qui se sont produit dans l’écologie de la Béringie et de nombreuses autres régions du globe au cours des derniers millénaires du Tardiglaciaire (Lundelius 1989).
Contexte chronologique
Comme nous l’avons vu, les données sédimentologiques, palynologiques et paléontologiques nous permettent de situer le remplissage des grottes le long d’une échelle chronologique qui recouvre très certainement l’Holocène (humus à cailloutis) et la fin du Pléistocène supérieur, c’est-à-dire, le Tardiglaciaire (lœss). Quelques datations au 14C obtenues en début de recherche (Cinq-Mars 1979; Morlan et Cinq-Mars 1982) ont confirmé cette appréciation chrono-stratigraphique pour le moins floue et y ont même apporté un avant-goût de précision. Nous avons ainsi pu dater un épisode de feu de forêt, nécessairement Holocène, dans des sédiments supérieurs (cryoturbés) de la grotte I. Nous avons également obtenu d’un fémur de cheval, provenant du sommet du lœss inférieur de la grotte I, un âge de 12,900 BP et, d’une omoplate de mammouth trouvée dans le lœss inférieur de la Grotte II où ont été identifiés les signaux polliniques de la toundra herbacée, un âge de 15,500 BP.
Une nouvelle série de datations au 14C obtenues récemment à partir de spécimens de la mégafaune (Note 5; Cinq-Mars et Nelson 1989) provenant des trois grottes nous autorise à mettre de l’avant une version chrono-stratigraphique un peu plus circonstanciée qui éclaire d’un jour nouveau certains problèmes relatifs à la paléontologie, à la paléœcologie et à l’archéoéogie.
Par exemple, certains de ces résultats peuvent nous aider à identifier ou préciser les périodes d’extinction et d’extirpation et, par retroaction, nous fournir de précieux renseignement concernant l’évolution du milieu. Le cas du saïga est, en ce sens, particulièrement instructif. Les exigences écologiques de cet animal (Vereshchagin et Baryshnikov 1982) en font un indicateur privilégié de milieu xérothermique (de type steppe semi-aride) relativement riche en espèces herbacées et caractérisé, en hiver, par de faibles accumulations de neige. Jusqu’à récemment, on datait sa présence en Béringie orientale d’avant le P1éniglaciaire (Harington 1980; Matthews 1982). Sa découverte dans les sédiments de la grotte III indique que des populations de saïga, vivant évidemment dans un milieu qui ne leur était pas entièrement défavorable, ont perduré à l’extrême-orient de la Béringie jusqu’à environ 13,400 BP. En fait, le chevauchement qui existe entre cette date et l’âge de la zone (palynologique) à bouleau, indicatrice, comme nous l’avons vu, d’un important changement écologique, signifie probablement que le saïga du Poisson-Bleu est un des derniers à avoir subsisté dans ces régions.
En plus d’apporter une confirmation évidente à la dichotomie lœss/humus à cailloutis et au fait que celle-ci date effectivement d’il y a environ 10,000 ans, une bonne partie de ces résultats chronométriques ajoute à notre chrono-stratigraphie une amplitude que nous ne lui soupçonnions pas. En effet, comme nous le verrons plus loin, certains des résultats indiquent que l’histoire du remplissage remonte jusqu’à environ 25,000 ans BP. Bien plus, ils servent à démontrer que la « faune à mammouth » mentionnée plus haut a constitué entre environ 17,000 et 25,000 BP, et sous une physionomie qui reste à déterminer avec précision, un é1ément essentiel du biotope pléniglaciaire de la Béringie orientale. Sans élaborer plus avant sur ces données qui servent à éclairer une importante controverse béringienne (Note 6), nous nous contenterons de souligner que cette démonstration de la viabilité du milieu pléniglaciaire est d’une importance capitale pour ce qui est de notre perception de certains aspects des réalités archéologiques béringiennes.
La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 3
LES TÉMOINS CULTURELS
À l’instar de nombreux gisements béringiens ayant fourni des matériaux archéologiques très anciens, les témoignages culturels provenant des grottes du Poisson-Bleu sont caractérisés par une rareté pour le moins exaspérante. Ils consistent en matériel lithique, lui-même divisible en trois catégories, en traces de décarnisation que l’on retrouve sur certains éléments de la mégafaune, ainsi qu’en quelques représentants de ce que l’on pourrait qualifier d’industrie de l’os peu élaborée. Sauf de rares exceptions (Note 7) , ils proviennent presque tous de la couche de lœss des grottes I et II où nous les trouvons dans des contextes dont la lecture, encore une fois, n’est pas toujours aisée.
Matériel lithique
La première catégorie consiste en une série composée d’environ cent pièces. Elle inclut d’un côté, des objets façonnés (Fig. 4) tels que nucléi, microlames, burins d’angle sur troncature, chutes de burin, encoche, etc. et, de l’autre, divers éclats de taille ou de retouche dont certains ont pu être utilisés. Les matières premières sont surtout des cherts bleutés (quelquefois mouchetés) ou, plus rarement, noirs, à grain très fin et dont les sources exactes, si elles nous sont encore inconnues, ne sont certainement pas régionales (Note 8). Jusqu’ici, de tels objets n’ont été récoltés que dans les sédiments des grottes I et II.

Ils proviennent en plus grande partie de l’intérieur et de l’entrée de la grotte II, à la base aussi bien qu’à la limite supérieure de cette zone (inférieure) du lœss qui est la plus riche en vestiges fauniques. Certaines des caractéristiques technologiques que présentent les burins (tous burins d’angle sur troncature) nous portent à croire qu’une bonne partie de l’ensemble représente peut-être un épisode spécifique d’utilisation de ce porche ou, encore, des visites consé ;cutives d’un groupe en particulier. Même si leur contexte ne nous permet pas de les situer chrono-stratigraphiquement avec précision, nous savons qu’ils datent de longtemps avant la fin de la période de mise en place du lœss, c’est-à-dire d’avant 10,000 ans BP. Leur provenance et leur association relative avec des restes d’espèces disparues nous portent à croire qu’ils ont pu être incorporés dans le gisement vers 12,000 BP et même avant.
Quelques outils, incluant une chute de burin, des fragments de microlames ainsi que des éclats (dont un, de retouche bifaciale) ont aussi été récoltés à l’intérieur de la grotte I, dans un contexte stratigraphique un peu plus précis (Cinq-Mars 1979). II semble, en effet, que certaines de ces pièces recouvrent, chronologiquement, la première moitié de la période caractérisée par la zone « à bouleau ». D’autres, cependant, dénotent des âges un peu plus récents (possiblement entre 10,000 et 12,000 BP) ainsi que plus anciens (avant 13,500 BP).
La deuxième catégorie est composée de plusieurs dizaines de petits éclats détritiques résultant de la taille, de la retouche ou, encore, de l’utilisation d’outils en chert (Note 9). Si petits soient-ils, ils possèdent toutes les caractéristiques morphologiques des éclats ou esquilles plus larges et se retrouvent en quantités variables dans tous les gisements archéologiques où ont été taillés ou utilisés des outils en pierre. Ceux qui proviennent du remplissage éolien des grottes du Poisson-Bleu, se démarquent bien, quant à la matière première, des autres particules rocheuses de dimensions semblables. Bien qu’ils aient été trouvés dans les sédiments des trois grottes, ce sont ceux de la grotte I qui nous fournissent le plus d’information. Leur mode de répartition à l’intérieur de la grotte chevauche celui que nous avons déjà décrit pour l’outillage. S’y ajoute aussi une composante suggestive de sédimentation culturelle dans cette partie du lœss qui présente une signature pollinique caractéristique de la toundra herbacée et dont l’âge, nous l’avons vu, recouvre la période allant de 25,000 à 13,500 BP.
Notre troisième catégorie comprend de nombreux petits galets roulés qui, pour la plupart, ont été récoltés à la base du lœss, en contact ou presque avec la roche-mère et, le plus souvent, près de l’entrée ou à l’intérieur des porches. Comme ils sont tous de nature clairement allochtone, nous avons cru, jusqu’à tout récemment (Morlan et Cinq-Mars 1982: Fig. 9), que leur présence dans le lœss ne pouvait s’expliquer que par le biais d’une action animale ou, dans le cas des plus gros spécimens, possiblement culturelle. Comme dans le cas des grottes, il s’avère que leur origine s’explique plus correctement dans le cadre de l’évolution d’un réseau karstique. Nous avons cependant choisi de conserver cette catégorie afin de pouvoir éventuellement y intégrer quelques uns des plus gros qui ont peut-être, après tout, servi de percuteurs.
Traces de dépeçage et de décarnisation
Malgré les difficultés inhérentes à toute tentative de décodage taphonomique, nous avons pu extraire de cet immense palimpseste (Binford 1981: 9) que forment, avec d’autres vestiges, les restes fauniques du Poisson-Bleu, des signes complémentaires de présence humaine.
Ces traces consistent en une variété d’entailles, d’incisions, de râclures, de hachures et de stries qui résultent d’activités intentionnelles de dépeçage et de décarnisation des animaux. Elles entament de façon plus ou moins superficielle, en divers endroits, les parois externes des ossements (Morlan et Cinq-Mars 1982: Fig. 10) et ont été effectuées avec des outils en pierre. Ajoutons que nous faisons, ici, référence à des signaux indéniablement culturels et non pas aux nombreuses imitations dont sont capables les carnivores, les rongeurs, divers processus géologiques ainsi que certains fouilleurs (Note 10). Jusqu’à présent, nous croyons avoir pu en identifier des exemples sur de nombreux éléments du squelette de presque toutes les espèces mégafauniques, à l’exclusion, peut-être, du loup, de l’orignal, du wapiti et du saïga. Presque tous proviennent des grottes I et II.
Ce type de données nous permet également d’accéder à une meilleure définition chronologique du contenu culturel du remplissage. Ainsi, comme nous n’avons pas trouvé de pareilles traces sur les restes fauniques provenant de l’humus à cailloutis, il est clair que le phénomène et ses causes sont d’âge pléistocène. Ceci est confirmé par certaines des dates 14C mentionnées plus haut et obtenues à partir de spécimens porteurs de ces modifications. Ces déterminations suggèrent que des activités culturelles reliées à l’exploitation de la faune du Poisson-Bleu se sont produites, de façon sporadique, entre environ 25,000 BP et 10,000 BP.
Industrie de l’os
Les exemples de cette industrie sont peu nombreux et peuvent être répartis en deux groupes. Le premier inclut des outils traditionnels un tant soit peu façonnés, intentionnellement ou par l’usage. Nous en avons trouvé des exemples dans les grottes I et II. Le deuxième groupe comprend des ossements taillés par percussion. Ils sont peu nombreux et proviennent de la grotte II.
Dans le premier cas, il s’agit surtout de quelques os longs qui, après avoir été fendus dans le sens de la longueur, ont peut-être été utilisés comme queursoirs, pour le traitement des peaux. L’un d’eux, en particulier, façonné à partir d’un tibia de caribou, porte des traces d’aménagement qui semblent résulter d’un travail de rabotage au burin; il présente, également, un poli très accentué, situé aux marges d’une cassure (d’utilisation ?) distale (Morlan et Cinq-Mars 1982: Fig.9). Cet objet a été récolté à l’extérieur de la grotte II, dans la zone inférieure du lœss. Il a aussi été daté à 24,820 BP, ce qui nous permet de mieux saisir non seulement l’amplitude chronologique du dépôt, comme nous l’avons noté précédemment, mais aussi l’amplitude chronologique des manifestations culturelles. Autrement dit, nous croyons pouvoir ajouter cet outil à une liste grandissante de données qui, bien qu’éparses, servent à démontrer que des populations humaines furent en mesure d’exploiter les ressources de la région, au Pléniglaciaire et même avant.

Il en va ainsi pour le deuxième groupe qui n’est représenté que par deux objets en os produits par percussion. Il s’agit d’un éclat (Note 11) et de son nucléus (Fig. 5) qui proviennent nécessairement du même os long d’un mammouth et qui ont été obtenus à partir d’une chaîne ou séquence opératoire relativement complexe que nous tenterons de schématiser comme suit:
- la matière première, c’est-à-dire, l’os long d’un mammouth, a d’abord été réduite à sa portion congrue qui comprend l’épiphyse et une partie attenante de la diaphyse;
- on a alors aménagé, à l’extrémité diaphysaire de ce bloc d’os ce que l’on pourrait qualifier de plan de frappe assez fruste;
- à partir de ce plan de frappe, on a ensuite détaché par percussion, de la face corticale du segment diaphysaire, une série de trois éclats dont les longueurs minimales varient entre sept et dix centimètres;
- enfin, un de ces éclats, le plus long, a été repris par taille ou retouche bifaciale et réduit en diagonale à partir de son extrémité proximale, de plus du tiers de ses dimensions originales.
Ces deux derniers témoins ont été trouvés à la base du lœss inférieur, dans des secteurs adjacents du remplissage situé en aval de l’entrée de la grotte II. Les deux dates qu’ils nous ont fournies révèlent un âge moyen, pléniglaciaire, d’environ 23,500 ans.
La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 4
INTERPRÉTATION ET CONCLUSION
Ces bribes d’information évoquent une images inévitablement cryptique et parcellaire, mais néanmoins cohérente, de quelques uns des phénomènes paléoécologiques et culturels qui ont marqué les confins orientaux de la Béringie au cours du Pléniglaciaire et du Tardiglaciaire. Elles ne peuvent atteindre, cependant, leur pleine signification qu’en étant confrontées à des ensembles plus vastes de données émanant d’un côté, de la Béringie dans son entier et, de l’autre, des régions plus méridionales de notre hémisphère. Cette mise en perspective ne pouvant se concevoir qu’à long terme et dans un autre cadre, nous nous contenterons d’en souligner ici quelques uns des éléments et des repères archéologiques les plus importants.
Considérations régionales et béringiennes
Pendant les 25 millénaires qu’embrasse le remplissage des grottes du Poisson-Bleu, les rares témoignages culturels que nous avons pu observés sont nettement limités à la tranche pléistocène durant laquelle ils se manifestent de façon sporadique et, le plus souvent diffuse, pendant environ 15,000 ans. Au cours de cette période, nous pouvons noter la présence de deux groupes de données dont la signification dépasse celui de la simple présence humaine.
Mentionnons, en premier lieu, la série lithique representative d’une industrie à microlames et burins. Les outils qu’elle contient peuvent être interprétés comme correspondant à quelques uns des éléments constitutifs du faciès régional d’un techno-complexe qui semble se manifester sur l’ensemble de la Béringie à la fin du Pléistocène (Note 12) et dont des exemples divers perdurent, dans certaines régions de la Béringie, pendant une partie de l’Holocène. Des traces en ont été trouvées en de nombreux sites du Yukon et de l’Alaska où elles forment une partie importante de ce que subsument des concepts tels que le complexe Denali (West 1967), la tradition American Paleoarctic (Anderson 1968; Dumond 1977) et la tradition Beringian (West 1981). Dans la plupart des cas, ce type de matériel se manifeste de concert avec des formes technologiques tels que des bifaces, des racloirs, des grattoirs, et des lames. Selon les données issues de divers gisements d’Alaska et, plus spécialement, de la région de la rivière Nenana où il est présentement étudié de façon détaillée, la présence de ce techno-complexe sur la scène béringienne orientale semble déjà bien établie autour de 10,500 BP et cela, dans la foulée d’un complexe archéologique antérieur nommé Nenana. Ce dernier, qui date d’environ 12,000 ans BP et qui est caractérisé par des pointes bifaciales ainsi que par une absence de microlames, a récemment été décrit comme faisant partie d’une lignée clovisienne sensu lato (Powers et Hoffecker 1989).
Cette série culturelle est considérée par les chercheurs que nous venons de citer comme une expression, peut-être tardive (Note 13), des premières explorations humaines en Béringie orientale. Ce qu’apportent les données du Poisson-Bleu, dans ce contexte, ne participe pas tellement du domaine de la caractérisation culturelle mais bien, plutôt, de celui de notre perception de la chronologie des événements. En effet, et ce, malgré la petitesse des échantillons et leur flou stratigraphique, elles suggèrent que des porteurs d’une industrie incorporant microlames et burins (et certainement plus) avaient atteint l’extrême est de la Béringie orientale bien avant le dernier millénaire du Tardiglaciaire.
Il en va de même pour l’autre groupe de données qui, dans la séquence du Poisson-Bleu, se situe au début du Pléniglaciaire, entre 23,000 et 25,000 ans BP. Nous faisons surtout référence à ces objets en os de mammouth taillés par percussion (éclat et nucléus), témoins osseux qui, dans ce cas-ci, fournissent un apport aussi bien dans le domaine de la caractérisation technologique que dans celui de la chronologie.
Le contexte particulier dans lequel ont été trouvés ces objets, aux grottes du Poisson-Bleu, nous permet de rejeter d’emblée une série d’objections ou de contre-hypothèses (Note 14; Cinq-Mars et Morlan 1989) mises de l’avant au cours des dernières années et dont le but était de nier à de semblables vestiges osseux provenant du bassin lacustre d’ Old Crow, une acception culturelle (Morlan 1980 , 1984). En effet, maints exemplaires d’ossements de mammouth recueillis dans les gisements fossilifères du bassin d’Old Crow dénotent des caractéristiques en tous points analogues à celles qui constituent l’essentiel de la chaîne opératoire que nous avons décrite plus haut. La répétition systématique d’une telle série de gestes ne pouvant certainement pas être le résultat d’aléas environnementaux, il nous faut considérer les vestiges qui en témoignent comme étant, sinon des outils, tout au moins des produits ou des sous-produits d’activités culturelles.
Cette relation évidente qui existe entre les données (en place) du Poisson-Bleu et celles (hors-contexte) du bassin d’Old Crow indiquent clairement que notre région, c’est-à-dire le bassin de la rivière Porcupine et les hautes terres environnantes, a été le théâtre d’une longue série de manifestations culturelles dont les plus anciennes, selon les données du bassin d’Old Crow, semblent apparaître autour de 40,000 ans BP (Morlan, et al. 1989). En d’autres termes, même s’il nous reste à en découvrir des exemples en Alaska et en Sibérie, il appert que les premiers signes décelables d’une présence humaine en Béringie remontent à l’interstade wisconsinien.
Considérations hémisphériques
Il découle de cette situation que la génération d’hypothèses concernant l’antiquité des premières explorations humaines dans l’ensemble de notre hémisphère, du débouché de la Béringie à la Tierra del Fuego, n’a pas à être limitée ou marquée par les seules contraintes écologiques et géographiques qui caractérisent le Tardiglaciaire béringien et qui servent, avec d’autres facteurs, à démontrer la primauté clovisienne en Amérique du Nord (Note 15).
S’il est vrai que ces dernières font partie des paramètres dont il faut tenir compte dans l’examen de la nature des liens qui ont dû exister, par exemple, entre le complexe Nenana et les cultures clovisiennes, elles ne tiennent certainement plus dans un contexte interstadial. La « chronologie courte » mentionnée précédemment pourrait ainsi n’être qu’un segment (parmi d’autres) d’une courbe ou d’une trajectoire espace-temps très complexe et de plus grande amplitude sur laquelle viendraient se greffer ces vestiges archéologiques trouvés plus au sud et que d’aucuns considèrent comme pré-clovisiens ou, pour utiliser la terminologie proposée par Meltzer (1989), « pré-12,000 BP » (Note 16).
Par ailleurs, il se peut que nos données les plus anciennes ne soient que le reflet d’une colonisation larvée qui, pour des raisons que l’on ignore, n’a jamais dépassé le seuil oriental de la Béringie et, qui plus est, ne présente aucun rapport de continuité avec les manifestations ultérieures. Il peut s’agir, au contraire, d’une longue pose, d’un temps d’arrêt au cours duquel se sont développés, chez des populations qu’il nous reste à identifier, ces modes d’adaptation qui facilitèrent à quelques unes d’entre elles, plus tard et très certainement au Tardiglaciaire, le passage vers le sud.
En conclusion, nous nous limiterons à souligner que les données du Poisson-Bleu nous permettent de mieux appréhender les limites du cadre chronologique dans lequel doivent être formulées les hypothèses concernant le peuplement initial de l’Amérique et ce, que ces dernières soient de nature strictement archéologique ou basées, en plus, sur les données de la linguistique et de la biologie (Greenberg et al. 1986). Enfin, ces données conduisent à la réalisation que l’espace béringien doit être perçu, non pas comme un simple lieu de passage entre deux pôles continentaux, ainsi que l’a déjà noté Workman (1980), mais bien, plutôt, comme un important creuset biologique et culturel de longue durée qui mérite d’être étudié comme tel si l’on veut, qu’un jour, il apporte sa juste contribution à notre compréhension de la suite des évènements dans une Amérique en voie de peuplement.
La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 5
BIBLIOGRAPHIE
- Adovasio, J.M. and R.C. Carlisle.
1986 « Pennsylvania Pioneers », Natural History Vol.95, pp. 21-27. - Anderson, D.D.
1968 « A Stone Age Campsite at the Gateway to America », Scientific American Vol. 218, pp. 24-33. - Binford, L.R.
1981 Bones. Ancient Men and Modern Myths. (New York, Academic Press). - Cinq-Mars J.
1979 « Bluefish Cave 1: A Late Pleistocene Eastern Beringian Cave Deposit in the Northern Yukon », Canadian Journal of Archaeology No. 3, pp. 1-32. - 1982 « Les grottes du Poisson-Bleu », Geos Vol. 11, pp. 19-21.
- Cinq-Mars J. and B. Lauriol
1985 « Le karst de Tsi-it-toh-choh: notes préliminaires sur quelques phénomènes karstiques du Yukon septentrional, Canada, Annales de la Société Géoloqique de Belgique T. 108, pp. 185-195. - Cinq-Mars, J. and R.E. Morlan.
1989 « The Peopling of the Americas as Seen from Northern Yukon Territory », (article submitted for publication in the proceedings of the World Summit Conference on the Peopling of the New World, Orono, Maine, May 1989). - Cinq-Mars, J. and E. Nelson.
1989 « AMS bone dates from the Bluefish Caves, northern Yukon, Canada », (article en préparation, 1989). - Cwynar, L. and J.C. Ritchie.
1980 « Arctic steppe-tundra: A Yukon perspective », Science Vol. 208, pp. 1375-1377. - Dillehay, T.D. and M. Collins.
1988 « Early Cultural Evidence from Monte Verde in Chile », Nature Vol. 332, pp. 150-152. - Dolitsky, A.B.
1985 « Siberian Paleolithic Archaeology: Approaches and Analytic Methods », Current Anthropology Vol. 26, pp. 361-378. Dumond, D.E., 1977 The Eskimos and Aleuts. (London, Thames and Hudson). - Fladmark, K.R.
1979 « Routes: Alternative Migration Corridors for Early Man in North America », American Antiquity Vol. 44, pp. 55-69. - 1982 « Microdebitage analysis: Initial Considerations », Journal of Archaeological Science Vol. 9, pp. 205-220. Greenberg, J.H., C.G. Turner, II and S.L. Zegura.
- 1986 « The Settlement of the Americas: A Comparison of the Linguistic, Dental, and Genetic Evidence », Current Anthropology Vol. 27, pp. 477-497.
- Gruhn, R.
1988 « Linguistic Evidence in Support of the Coastal Route of Earliest Entry into the New World », Man (N.S.) Vol. 23, pp. 77-100. - Gruhn, R. and A.L. Bryan.
1984 « The Record of Pleistocene Megafaunal Extinctions at Taima-Taima, Northern Venezuela ». In: Quaternary Extinctions: A Prehistoric Revolution, ed. P.S. Martin and R.G. Klein. (Tucson, University of Arizona Press) pp. 128-137. - Guidon, N. and G. Delibrias.
1986 « Carbon-14 Dates Point to Man in the Americas 32,000 Years Ago », Nature Vol. 321, pp. 769-771. - Guthrie, R.D.
1982 « Mammals of the Mammoth Steppe as Paleoenvironmental Indicators. » In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 307-326. - 1985 « Woolly Arguments Against the Mammoth Steppe – A New Look at the Palynological Data », Quarterly Review of Archaeology Vol. 6.
- Harington, C.R.
1980 « Radiocarbon Dates on Some Quaternary Mammals and Artifacts from Northern North America », Arctic Vol. 33, pp. 815-832. - 1989 « Pleistocene Vertebrate Localities in the Yukon. » In: Late Cenozoic History of the Interior Basins of Alaska and the Yukon, ed. L.D. Carter, T.D. Hamilton and J.P. Galloway (U.S. Geological Survey Circular 10269) pp. 93-98.
- Haynes, C.V.
1982 « Were Clovis Progenitors in Beringia? » In: Paleœcology of Beringa, édit. par D.M. Hopkins et al. (New York, Academic Press) pp. 383-398. - Hopkins, D.M.
1959 « Cenozoic History of the Bering Land Bridge (Alaska) », Science Vol. 129, pp.1519-1528. - Hopkins, D.M. (Éditeur)
1967 The Bering Land Bridge. (Stanford, Stanford University Press). - Hopkins D.M., J.V. Matthews, Jr., C.E. Schweger and S. Young (Éditeurs) 1982 Paleoecology of Beringia. (New York, Academic Press).
- Hughes O.L.
1972 « Surficial geology of northern Yukon Territory and northwestern District of Mackenzie », Geological Survey of Canada Paper 69-36. - Hultén, E.
1968 Flora of Alaska and neighboring territories. (Stanford University Press). - Irving, W.N., N.A. Jopling and B. Beebe.
1986 « Indications of Pre-Sangamon Humans near Old Crow, Yukon, Canada. » In: New Evidence for the Pleistocene Peopling of the Americas, ed. A.L Bryan (Orono, Center for the Study of Early Man) pp. 49-63. - Ives, J.W., A.B. Beaudoin and M.P.R. Magne.
1989 « Evaluating the Role of a Western Corridor in the Peopling of the Americas », (comm. prép. pour la Circum-Pacific Prehistory Conference, Seattle, Washington, August 1989). - Lauriol B., D.C. Ford and J. Cinq-Mars.
1989 « Landscape development from caves and speleothem data. Preliminary evidence from northern Yukon, Canada », (article submitted to Journal canadien des sciences de la terre). - Lumley, H. and M. de, M.C. Beltralo, Y. Yokoyama, J. Labeyrie, J. Danon, G. Delibrias, C. Falguères and J.L. Bischoff.
1987 « Présence d’outils taillés associés à une faune quaternaire datée du Pléistocène moyen dans la Toca da Esperanga, région de Central, état de Bahia, Brésil », L’Anthropologie T. 91, pp.917-942. - Lundelius, E.L., Jr.
1989 « The Implications of Disharmonious Assemblages for Pleistocene Extinctions », Journal of Archaeoloaical Sciences Vol. 16, pp. 407-417. - Martin, P.S.
1982 « The Pattern and Meaning of Holarctic Mammoth Extinction. » In: Paleoecology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 399-408. - Matthews, J.V., Jr.
1982 « East Beringia During Late Wisconsin Time: A Review of the Biotic Evidence. » In: Paleœcology of Beringia, ed. D.M. Hopkins et al. (New York, Academic Press) pp. 127-150. - Mochanov, Yu.
1978 « The Paleolithic of NE Asia and the problem of the first peopling of America. » In: Early Man in America from a Circum-Pacific perspective, édit. par A.L. Bryan (University of Alberta Department of Anthropology Occasional Paper 1) p. 67. - 1980 « Early migrations to America in the light of a study of the Dyuktai Paleolithic culture in Northeast Asia. » In: Early Native Americans, ed. D.L. Brownman (La Haye, Mouton) pp. 119-131.
- Morlan, R.E.
1980 Taphonomy and archaeology in the Upper Pleistocene of the northern Yukon Territory: a glimpse of the peopling of the New World. (Ottawa, Musée national de l’Homme, série Mercure, Commission archéologique du Canada, dossier no. 94). - Morlan, R.E.
1984 « Toward the Definition of Criteria for the Recognition of Artificial Bone Alterations. » Quaternary Research Vol. 22, pp. 160-171. - Morlan, R.E.
1989 « Paleœcological implications of Late Pleistocene and Holocene microtine rodents from the Bluefish Caves, northern Yukon Territory », Canadian Journal of Earth Sciences Vol. 26, pp. 149-156. - Morlan, R.E. and J. Cinq-Mars.
1982 « Ancient Beringians: Human Occupation in the Late Pleistocene of Alaska and the Yukon Territory. » In: Paleœcology of Beringia, édit. par D.M. Hopkins et al. (New York, Academic Press) pp. 353-381. - Morlan, R.E., D.E. Nelson, T.A. Brown, J.S. Vogel and J.R. Southon.
1989 « Accelerator Mass Spectrometry Dates on Bones from the Old Crow Basin », Canadian Journal of Archaeology Vol. 14, pp.75-92. - Norris, D.K.
1985 Stratigraphy of Devonian Outcrop Belts in Northern Yukon Territory and Northwestern District of Mackenzie (operation Porcupine area). (Ottawa, Mémoire 410, Commission géologique du Canada). - Powers, W.R. and J.F. Hoffecker.
1989 « Late Pleistocene Settlement in the Nenana Valley, Central Alaska », American Antiquity Vol. 54, pp. 263-287. - Ritchie, J.C.
1984 Past and Present Vegetation of the Far Northwest of Canada. (Toronto, University of Toronto Press). - Ritchie J.C., J. Cinq-Mars and L. Cwynar.
1982 « L’environnement tardiglaciaire du Yukon septentrional, Canada », Géographie physique et Quaternaire Vol. XXXVI, pp. 241-250. - Ritchie, J.C. and L. Cwynar.
1982 « The Late Quaternary Vegetation of the North Yukon. » In: : Paleœcology of Beringia, édit. par D.M. Hopkins et al. (New York, Academic Press) pp. 113-126. - Roberge, J., B. Lauriol, P. Thibaudeau and J. Cinq-Mars.
1986 « Caractère des karsts arctiques du Yukon septentrional », Comptes rendus du IXème Congrès international de spéléologie (Barcelone), Vol. 1, pp. 164-167. - Schweger, C.E., J.V. Mathews, Jr., D.M. Hopkins and S.B. Young (Éditeurs)
1982 « Paleoecology of Beringia – A Synthesis. » In: Paleœcology of Beringia, édit. par D.M. Hopkins et al. (New York, Academic Press) pp. 425-444. - Shipman, P.
1981 « Applications of Scanning Electron Microscopy to Taphonomic Problems. » In: The Research Potential of Anthropological Museum Collections, ed. A.-M. Cantwell, J. Griffin and N.A. Rothschild (New York, Annals of the New York Academy of Sciences 376), pp. 357-386. - Vereshchagin, N.K. and O.F. Baryshnikov.
1982 « Paleoecology of the Mammoth Fauna in the Eurasian Arctic. » In: Paleœcology of Beringia, édit. par D.M. Hopkins et al. (New York, Academic Press), pp. 267-279. - West, F.H.
1967 « The Donnelly Ridge Site and the Definition of an Early Core and Blade Complex in Central Alaska », American Antiquity Vol. 32, pp. 300-382. - West, F.H.
1981 The archaeology of Beringia. (New York, Columbia University Press). - Workman, W.B.
1980 « Holocene Peopling of the New World: Implications of the Arctic and Subarctic Data. » In: The Ice-Free Corridor and Peopling of the New World, , édit. par N.W. Rutter and C.E. Schweger (Revue canadienne d’Anthropologie , Vol. 1), pp. 129-139. - Yi, S. and G. Clark.
1985 « The ‘Dyuktai Culture’ and New World Origins », Current Anthropology Vol. 26, pp. 1-20.
La place des Grottes du Poisson-Bleu dans la préhistoire béringienne – Page 6
NOTES
- Nous faisons référence aux options béringiennes stricto sensu ainsi qu’au concept de « couloir du Mackenzie » (Ives et al. 1989), sans pour autant nier la valeur de l’hypothèse « côtière » mise de l’avant par Fladmark (1979) et reprise par Gruhn (1988).
- Les grottes ont été découvertes accidentellement en 1975 et des sondages et des fouilles y ont été effectués, de façon intermittente, entre 1977 et 1987 (Cinq-Mars 1979 et 1982; Morlan et Cinq-Mars 1982).
- L’analyse ayant démontré que le lœss ne provient pas du paysage calcaire environnant (C. Tarnocai, comm. pers.), il nous faut chercher son origine dans les sédiments mis au jour au cours des cycles d’exondation des grands bassins lacustres (lacs glaciaires Bluefish et Old Crow) situés plus au nord.
- Ce contraste est aussi bien illustré par les résultats de l’étude d’une partie des microtinés des Grottes I et II, récemment effectuée par R. Morlan (1989).
- Toutes les datations de spécimens fauniques proviennent de la fraction collagène des ossements et, à part deux exceptions, ont été obtenues grâce à l’analyse spectrométrique de masse par accélérateur effectuée par R.M. Brown (Énergie atomique du Canada) et, plus particulièrement, par E. Nelson (Université Simon Fraser).
- Il s’agit d’une controverse concernant la nature exacte et la viabilité relative du milieu béringien au cours du Pléniglaciaire supérieur, l’ensemble du problème ayant été qualifié de productivity paradox (Schweger et al. 1982). Pour d’aucuns (Cwynar et Ritchie 1980), ces régions n’auraient pu supporter, au mieux, qu’un biome très appauvri; selon d’autres, au contraire, le milieu aurait été biologiquement très riche (Guthrie 1985; Matthews 1982).
- Deux objets (un nucléus et un fragment de microlame) ont été découverts dans l’humus à cailloutis de la grotte II, dans des contextes vraisemblablement remaniés. Une des crêtes de l’affleurement calcaire du Poisson-Bleu a également fourni, en surface, quelques petits éclats ainsi qu’une microlame.
- Les calcaires du Poisson-Bleu ne contiennent pas de chert. Les quelques exemples de ce matériaux trouvés dans la région (dans quelques affleurements rocheux situés à l’est de grottes ou sous forme de galets roulés dans les lits de rivières) sont de très mauvaise qualité. Par ailleurs, nous savons que des cherts semblables à ceux des grottes I et II sont fréquemment trouvés dans les nombreux gisements archéologiques situés plus de cent kilomètres au nord, le long des contreforts de la Cordillère (chaînons Brooks, British et Barn).
- En général plus petits que 0.003 m, ces éclats sont recueillis lors du traitement des échantillons de sédiments (Cinq-Mars 1979; Fladmark 1982).
- Jusqu’à présent, l’identification de ces traces dont la lecture est souvent difficile a été faite à l’aide du microscope binoculaire. Nous entreprendrons sous peu un travail de verification au microscope à balayage électronique (MEB) en utilisant la démarche proposée par Shipman (1981).
- Mentionnons aussi que cet éclat participe de la catégorie précédente puisqu’il présente sur sa face dorsale, une courte incision causée par un tranchant.
- Ce techno-complexe est mieux connu, en Sibérie, sous le nom de culture ou tradition Diouktai (Mochanov 1978, 1980). Comme il est hors de notre propos de discuter de son origine, de son ancienneté et, même, de sa signification, nous nous contenterons de souligner que ces problèmes sont loins d’être résolus (Dolitsky 1985; Yi et Clark 1985). Néanmoins, selon ces auteurs, sa présence dans le nord-est de la Sibérie semble clairement attestée à la fin du Pléniglaciaire, autour de 18,000 ans BP, ce qui s’accorde bien avec l’âge que nous croyons pouvoir lui assigner, en Béringie orientale, à partir des données du Poisson-Bleu.
- La localisation des manifestations culturelles qui intéressent Powers et Hoffecker (1989) semble pouvoir s’expliquer par le fait que la vallée de la Nenana ainsi que certaines régions avoisinantes furent, à la fin du Tardiglaciaire, des zones-refuges où se retrouvèrent les derniers vestiges de la toundra herbacée et de la « faune à mammouth ». On peut donc supposer qu’y perdurèrent aussi, chez certains groupes humains, des modes d’adaptation et d’exploitation du milieu dont l’apprentissage était vraisemblablement pan-béringien et, nécessairement, plus ancien.
- Le lecteur pourra trouver dans un autre article (Cinq-Mars et Morlan 1989) une critique de ces contre-hypothèses qui s’appuient sur divers phénomènes naturels afin d’expliquer la présence d’ossements de mammouth taillés par percussion dans les gisements fossilifères de la rivière Old Crow. Notons, par ailleurs, que nous ne nions pas la justesse de ces contre-hypothèses lorsqu’il s’agit de mettre en lumière les causes de certains autres types de modification sur des ossements d’espèces diverses.
- Il s’agit, bien entendu, des thèses défendues par Paul Martin (1982) et Vance Haynes (1982).
- Sans nécessairement les accepter d’emblée, nous faisons référence à des gisements tels que Meadowcroft (Adovasio et Carlisle 198,6), Monte Verde (Dillehay et Collins 1988), Toca do Boqueirao da Pedra Furada (Guidon et Delibrias 1986) et Taima-Taima (Gruhn et Bryan 1984). Il est évident, par ailleurs, que contrairement à celui mis de l’avant par Irving et ses collègues (Irving et al. 1986), le cadre de référence chronologique béringien que nous avons tenté d’illustrer n’est pas encore en mesure de s’accommoder d’un gisement tel que la Toca da Esperança (de Lumley et al. 1987).